O pH refere-se a uma medida que indica se uma solução líquida é ácida (pH < 7, a 25 °C), neutra (pH = 7, a 25 °C), ou básica/alcalina (pH > 7, a 25º C). Uma solução neutra só tem o valor de pH = 7 a 25 °C, o que implica variações do valor medido conforme a temperatura.
Definição
pH é o símbolo para a grandeza físico-química 'potencial hidrogeniônico'. Essa grandeza indica a acidez, neutralidade ou alcalinidade de uma solução aquosa.
O termo pH foi introduzido, em 1909, pelo bioquímico dinamarquês Søren Peter Lauritz Sørensen (1868-1939) com o objetivo de facilitar seus trabalhos no controle de qualidade de cervejas (à época trabalhava no Laboratório Carlsberg, da cervejaria homônima). O "p" vem do alemão potenz, que significa poder de concentração, e o "H" é para o íon de hidrogênio (H+).
Às vezes é referido do latim pondus hydrogenii.
Matematicamente, o "p" equivale ao simétrico do logaritmo (cologaritmo) de base 10 da atividade dos íons a que se refere. Para íons H+
pOH
Do mesmo modo pode-se definir o pOH em relação à concentração de íons OH-. A partir da constante de dissociação da água que tem o valor de 10−14 à temperatura de 298 K (25 °C), pode-se determinar a relação entre pOH e pH. Assim, pela definição de Kw (produto iônico da água) tem-se a relação entre as duas atividades:
Kw = [H+] · [OH-]
Ao aplicar logaritmos, obtém-se a relação entre pH e pOH:
pKw = pH + pOH = 14
[editar] Cálculo de pH de algumas soluções aquosas
O valor de pH de uma solução pode ser estimado se for conhecida a concentração em íons H+. Apresentam-se em seguida vários exemplos:
[editar] Solução aquosa de ácido clorídrico (HCl) 0,1 mol L-1
Está é uma solução de um ácido forte, por isso, o HCl presente estará completamente ionizado. Como a concentração é de apenas 0,1 mol L−1, ele está suficientemente diluído para que os valores de sua atividade sejam próximos ao de sua concentração. Sendo assim, pode-se obter o pH pela expressão abaixo:
[H+] = 0,1 mol L−1
Então: pH = -log[0,1] = 1.
Alguns valores comuns de pH Substância pH
Ácido de bateria < 1,0
Suco gástrico 1,0 - 3,0
Sumo de limão 2,2 - 2,4
Refrigerante tipo cola 2,5
Vinagre 2,4-3,4
Sumo de laranja ou maçã 3,5
Cervejas 4,0 - 5,0
Café 5,0
Chá 5,5
Chuva ácida < 5,6
Saliva pacientes com câncer (cancro) 4,5 - 5,7
Leite 6,3 - 6,6
Água pura 7,0
Saliva humana 6,5 - 7,5
Sangue humano 7,35 - 7,45
Água do mar 8,0
Sabonete de mão 9,0 - 10,0
Amoníaco 11,5
"Água sanitária" 12,5
Hidróxido de sódio (soda cáustica) 13,5
[editar] Solução aquosa de hidróxido de sódio (NaOH) 0,1 mol L-1
Esta é uma solução de uma base forte, sendo assim, o NaOH presente está completamente dissociado. Como sua concentração é de apenas 0,1 mol L−1, ele está suficientemente diluído para que seu valor de atividade seja próximo ao da concentração. Sendo assim:
[OH-] = 0,1 mol L−1 Então: pOH = -log[0,1] = 1.
Pela relação entre pH e pOH, tem-se:
pH + pOH = 14
pH = 14-1 = 13
[editar] Solução aquosa de ácido fórmico (HCOOH) 0,1 mol L-1
Esta é uma solução de um ácido fraco, que por sua vez, não está completamente ionizado. Por isso deve-se determinar primeiro a concentração de H+.
Para ácidos fracos deve-se considerar a constante de dissociação do ácido (Ka):
Ka = [H+][HCOO-] / [HCOOH]
A constante de dissociação do ácido fórmico tem o valor de Ka = 1,6 × 10−4. Assim, considerando que [A-] é igual a x, [HA] há-de ser a parte que não se dissociou, ou seja 0,1-x. Se desprezarmos a ionização da água, concluímos que a única fonte de H+ é o ácido, assim [H+] = [A-]. Substituindo as variáveis obtém-se:
1.6\times 10^{-4} = \frac{x^2}{0.1-x}
A solução é:
[H+] = x = 3,9 × 10−3.
Através da definição de pH, obtém-se:
pH = -log[3,9 × 10−3] = 2,4
FONTE - wikipedia
segunda-feira, 19 de julho de 2010
domingo, 4 de julho de 2010
8° série - Ácidos e Bases
Ácidos e bases (também chamadas de álcalis) são costumeiramente lembrados como substâncias químicas perigosas, corrosivos capazes de dissolver metais como se fossem comprimidos efervecentes. Mas a presença dos ácidos e base na nossa vida cotidiana é bem mais ampla e menos agressiva do que se imagina.
Eles também são componentes usuais de refrigerantes, alimentos, remédios, produtos de higiene ou cosméticos. São ainda matérias primas indispensáveis em um vasto universo de aplicações industriais. A tal ponto que a produção de ácido sulfúrico e soda cáustica de um país chega a ser considerada um dos indicadores do seu nível de atividade econômica.
Definições de ácidos e bases
A definição mais tradicional dos ácidos e bases foi dada pelo cientista sueco Svante Arrhenius, que estabeleceu os ácidos como substâncias que - em solução aquosa - liberam íons positivos de hidrogênio (H+), enquanto as bases, também em solução aquosa, liberam hidroxilas, íons negativos OH-.
Outras definições de ácidos e bases
Uma outra definição para ácidos e bases foi dada pelo dinamarquês Johannes N. Bronsted e pelo inglês Thomas Lowry, independentemente, ficando conhecida como definição protônica. Segundo os dois, ácido é uma substância capaz de ceder um próton a uma reação, enquanto base é uma substância capaz de receber um próton.
A definição de Bronsted-Lowry é mais abrangente que a de Arrhenius, principalmente pelo fato de nem todas as substâncias que se comportam como bases liberarem uma hidroxila OH-, como é o caso da amônia (NH3). Além disso, a definição protônica não condiciona a definição de ácidos e básicos à dissolução em meio aquoso, como propunha a do químico sueco.
Bronsted e Lowry definiram ácidos e bases a partir dos prótons que liberavam e recebiam. Já o norte-americano Gilbert Newton Lewis se voltou para os elétrons ao desenvolver sua definição. De acordo com ela, ácidos são substâncias que, numa ligação química, podem receber pares eletrônicos, enquanto as bases são aquelas que cedem estes pares.
A definição de Lewis abrange as de Arrhenius e a definição protônica, que, entretanto, continuam válidas dentro de suas próprias abrangências.
Identificação dos ácidos e bases
Os ácidos possuem sabor azedo, como o encontrado nas frutas cítricas ricas no ácido de mesmo nome. Já as base tem gosto semelhante ao do sabão (sabor adstringente). Mas, felizmente, há modos mais eficazes e seguros de identificar ácidos e bases do que o paladar.
É possível medir a concentração de hidrogênio iônico em uma solução a partir de uma escala logarítmica inversa, que recebeu o nome de potencial hidrogeniônico, ou simplesmente, escala de pH.
Esta escala vai de zero a 14, sendo o pH 7 considerado neutro. Os valores menores que sete classificam a solução medida como ácida e os maiores que sete, como alcalinos (bases).
Para se medir o pH, usam-se combinações de substâncias indicadoras, como a fenolftaleína, que mudam de cor conforme a posição da substância testada na escala acima.
Também são usados instrumentos como os medidores de pH por eletrodo indicador, que mede as diferenças de potencial elétrico produzidas pelas concentrações de hidrogênio e indica o resultado dentro da escala de 0 a 14.
http://educacao.uol.com.br/quimica/ult1707u8.jhtm
Eles também são componentes usuais de refrigerantes, alimentos, remédios, produtos de higiene ou cosméticos. São ainda matérias primas indispensáveis em um vasto universo de aplicações industriais. A tal ponto que a produção de ácido sulfúrico e soda cáustica de um país chega a ser considerada um dos indicadores do seu nível de atividade econômica.
Definições de ácidos e bases
A definição mais tradicional dos ácidos e bases foi dada pelo cientista sueco Svante Arrhenius, que estabeleceu os ácidos como substâncias que - em solução aquosa - liberam íons positivos de hidrogênio (H+), enquanto as bases, também em solução aquosa, liberam hidroxilas, íons negativos OH-.
Outras definições de ácidos e bases
Uma outra definição para ácidos e bases foi dada pelo dinamarquês Johannes N. Bronsted e pelo inglês Thomas Lowry, independentemente, ficando conhecida como definição protônica. Segundo os dois, ácido é uma substância capaz de ceder um próton a uma reação, enquanto base é uma substância capaz de receber um próton.
A definição de Bronsted-Lowry é mais abrangente que a de Arrhenius, principalmente pelo fato de nem todas as substâncias que se comportam como bases liberarem uma hidroxila OH-, como é o caso da amônia (NH3). Além disso, a definição protônica não condiciona a definição de ácidos e básicos à dissolução em meio aquoso, como propunha a do químico sueco.
Bronsted e Lowry definiram ácidos e bases a partir dos prótons que liberavam e recebiam. Já o norte-americano Gilbert Newton Lewis se voltou para os elétrons ao desenvolver sua definição. De acordo com ela, ácidos são substâncias que, numa ligação química, podem receber pares eletrônicos, enquanto as bases são aquelas que cedem estes pares.
A definição de Lewis abrange as de Arrhenius e a definição protônica, que, entretanto, continuam válidas dentro de suas próprias abrangências.
Identificação dos ácidos e bases
Os ácidos possuem sabor azedo, como o encontrado nas frutas cítricas ricas no ácido de mesmo nome. Já as base tem gosto semelhante ao do sabão (sabor adstringente). Mas, felizmente, há modos mais eficazes e seguros de identificar ácidos e bases do que o paladar.
É possível medir a concentração de hidrogênio iônico em uma solução a partir de uma escala logarítmica inversa, que recebeu o nome de potencial hidrogeniônico, ou simplesmente, escala de pH.
Esta escala vai de zero a 14, sendo o pH 7 considerado neutro. Os valores menores que sete classificam a solução medida como ácida e os maiores que sete, como alcalinos (bases).
Para se medir o pH, usam-se combinações de substâncias indicadoras, como a fenolftaleína, que mudam de cor conforme a posição da substância testada na escala acima.
Também são usados instrumentos como os medidores de pH por eletrodo indicador, que mede as diferenças de potencial elétrico produzidas pelas concentrações de hidrogênio e indica o resultado dentro da escala de 0 a 14.
http://educacao.uol.com.br/quimica/ult1707u8.jhtm
7° série: SISTEMA LINFÁTICO

SISTEMA LINFÁTICO
Sistema paralelo ao circulatório, constituído por uma vasta rede de vasos semelhantes às veias (vasos linfáticos), que se distribuem por todo o corpo e recolhem o líquido tissular que não retornou aos capilares sangüíneos, filtrando-o e reconduzindo-o à circulação sangüínea.
É constituído pela linfa, vasos e órgãos linfáticos.
Os capilares linfáticos estão presentes em quase todos os tecidos do corpo. Capilares mais finos vão se unindo em vasos linfáticos maiores, que terminam em dois grandes dutos principais: o duto torácico (recebe a linfa procedente da parte inferior do corpo, do lado esquerdo da cabeça, do braço esquerdo e de partes do tórax) e o duto linfático (recebe a linfa procedente do lado direito da cabeça, do braço direito e de parte do tórax), que desembocam em veias próximas ao coração.
Linfa: líquido que circula pelos vasos linfáticos. Sua composição é semelhante à do sangue, mas não possui hemácias, apesar de conter glóbulos brancos dos quais 99% são linfócitos. No sangue os linfócitos representam cerca de 50% do total de glóbulos brancos.
Imagem: CRUZ, Daniel. O Corpo Humano. São Paulo, Ed. Ática, 2000.
Órgãos linfáticos: amígdalas (tonsilas), adenóides, baço, linfonodos ( nódulos linfáticos) e timo (tecido conjuntivo reticular linfóide: rico em linfócitos).
Amígdalas (tonsilas palatinas): produzem linfócitos.
Timo: órgão linfático mais desenvolvido no período prenatal, involui desde o nascimento até a puberdade.
Linfonodos ou nódulos linfáticos: órgãos linfáticos mais numerosos do organismo, cuja função é a de filtrar a linfa e eliminar corpos estranhos que ela possa conter, como vírus e bactérias. Nele ocorrem linfócitos, macrófagos e plasmócitos. A proliferação dessas células provocada pela presença de bactérias ou substâncias/organismos estranhos determina o aumento do tamanho dos gânglios, que se tornam dolorosos, formando a íngua.
Baço: órgão linfático, excluído da circulação linfática, interposto na circulação sangüínea e cuja drenagem venosa passa, obrigatoriamente, pelo fígado. Possui grande quantidade de macrófagos que, através da fagocitose, destroem micróbios, restos de tecido, substâncias estranhas, células do sangue em circulação já desgastadas como eritrócitos, leucócitos e plaquetas. Dessa forma, o baço “limpa” o sangue, funcionando como um filtro desse fluído tão essencial. O baço também tem participação na resposta imune, reagindo a agentes infecciosos. Inclusive, é considerado por alguns cientistas, um grande nódulo linfático.
Origem dos linfócitos: medula óssea (tecido conjuntivo reticular mielóide: precursor de todos os elementos figurados do sangue).
Linfócitos T – maturam-se no timo.
Linfócitos B – saem da medula já maduros.
Os linfócitos chegam aos órgãos linfáticos periféricos através do sangue e da linfa.
http://www.afh.bio.br/imune/linfa1.asp
6° série: PEIXES







OS PEIXES
"Vertebrados adaptados para viver apenas na água"
Adaptados exclusivamente ao ambiente aquático, os peixes compreendem cerca de 25.000 espécies, um número maior do que a soma de todas as espécies de vertebrados terrestres conhecidos.
Admite-se que os primeiros vertebrados surgiram há cerca de 500 milhões de anos nos mares e eram desprovidos de mandíbulas. Esses animais, chamados de ostracodermos, eram pequenos e viviam no fundo do mar, filtrando alimentos que se encontravam no lodo. Há cerca de 450 milhões de anos, a partir de alguns ostracodermos, surgiram os placodermos, vertebrados que possuíam mandíbulas, que permitiam a eles atuar como eficientes predadores. A lampreia descende dos antigos ostracodermos. Os peixes atuais e todos os demais vertebrados descendem dos placodermos.
Os peixes constituem a maioria dos vertebrados e todos têm, em comum, muitas características que os adaptaram à vida na água. Os peixes ancestrais não possuíam mandíbula, eram bentônicos e pertencentes à classe Agnatha. A maioria dos agnatos está extinta, mais a classe ainda é representada hoje em dia pelas lampreias e peixes-bruxa.
Com a evolução das mandíbulas e dos apêndices pares, os peixes tornam-se mais ativos e capazes de alimentarem-se de diferentes maneiras. Os peixes mandibulados atuais estão pados em duas classes: os tubarões e raias na classe Chondrichthyes, com esqueleto cartilaginoso, e as percas e outros peixes similares da classe Osteichthyes, que possuem um esqueleto ossificado pelo menos em parte. As características distintas das classes existentes são resumidas a seguir.
CLASSIFICAÇÃO E DIVERSIDADE
Os peixes são classificados em três grandes classes: Agnatha, Chondricthyes, Osteichthyes, abaixo:
CARACTERÍSTICAS DA CLASSE AGNATHA
As mandíbulas estão ausentes. As nadadeiras pares estão ausentes na maioria das espécies, as abas peitorais estavam presentes em algum formas extintas. As espécies primitivas tinham a pele revestida por formes escamas ósseas, que foram perdidas nas atuais. As partes mais internas do esqueleto são cartilaginosas nas formas atuais e parece que nas espécies extintas elas também não eram ossificadas. O notocórdio embrionário persiste nos adultos. Um olho pineal mediano e fotossensível está presente. As espécies atuais, como a maioria das extintas, apresentam uma narina única e mediana, localizada à frente do olho pineal. Sete ou mais aberturas branquiais estão presentes. A faringe é utilizada, na alimentação por filtração nas larvas e nos adultos das espécies que estão atualmente extintas, isto é, não são mais encontradas.
CARACTERÍSTICAS DA CLASSE CHONDRICHTHYES
As mandíbulas e as nadadeiras pares estão presentes. As escamas ósseas estão reduzidas a delgadas escamas placóides ou foram completamente perdidas. As partes mais internas do esqueleto são totalmente cartilaginosas. O olho pineal foi perdido. Eles são peixes compactos, sem pulmão ou bexiga natatória. Seus corpos são achatados no sentido ântero-central e a maioria das espécies continua com a cauda heterocerca primitiva. Suas narinas são pares. Os cinco pares de aberturas branquiais abrem-se independentemente na superfície corporal na maioria das espécies, ao contrário daquelas em que uma câmara branquial está recoberta por um opérculo. intestino é curto e a área superficial é aumentada por uma válvula espiral. Os machos possuem um clasper sobre a nadadeira pélvica, que transfere os espermatozóides para a fêmea. A fecundação é interna.
CARACTERÍSTICAS DA CLASSE OSTEICHTHYES
Em geral, as escamas ósseas estão presentes, mas as camadas superficiais primitivas de ganoína e cosmina foram perdidas na maioria das espécies atuais. As partes mais internas do esqueleto sempre apresentam alguma ossificação; na maioria das espécies, o esqueleto é completamente ossificado. O olho pineal continua presente nas espécies primitivas. Pulmões ou bexigas natatórias estão presentes, exceto em poucas espécies bentônicas, que os perderam secundariamente. Como não poderia deixar de ser nos peixes de corpos ágeis, a cauda tornou-se homocerca na maioria das espécies atuais. As aberturas branquiais se abrem numa câmara comum, coberta por um opérculo. A válvula espiral do intestino foi perdida em todas as espécies, exceto na maioria das primitivas. A área superficial é maior devido a um aumento do comprimento do intestino e cecos pilóricos. A maioria das espécies é ovípara e a fecundação é externa. Em algumas espécies vivíparas, nas quais a fecundação é interna, o órgão copulador do macho é uma parte modificada da nadadeira anal.
Os peixes estão bem adaptados à vida aquática. Eles são aerodinâmicos. Seu esqueleto não é tão pesado como o dos vertebrados terrestres. Os músculos segmentados e a cauda proporcionam o impulso para a locomoção e as nadadeiras, a estabilidade e a habilidade de manobras.
A estrutura dos órgãos dos sentidos permite a detecção de mudanças ocorridas na água. Seu coração bombeia apenas sangue venoso - através das brânquias. Uma língua muscular está ausente. Os peixes mais primitivos, que surgiram antes do período Cambriano superior, eram ostracodermos possuidores de armaduras resistentes e pertencentes à classe Agnatha. A maioria era dulcicola e alimentava-se de sedimentos, com a boca sem mandíbulas. Eles não apresentavam nadadeiras pares bem desenvolvidas e não eram peixes muito ativos Os únicos vertebrados agnatos ainda existentes são as lampreias e peixes-bruxa da ordem Ciclostomata. Eles também não possuem mandíbulas nem apêndices pares. As mandíbulas, que surgiram primeiro nos acantódios, evoluíram de um arco visceral alargado, o arco mandibular. Ossos dérmicos podem unir-se ao arco mandibular. Os acantódios possuíam mais de dois aparelhos de apêndices pares, que eram sustentados por espinhos. Os placodermos são uma classe de peixes primitivos extintos, muitos dos quais possuíam mandíbulas semelhantes a machadinhas.
Os peixes cartilaginosos da classe Chondrichthyes caracterizam-se por possuir pequenas escamas placóides, esqueleto sem osso, ausência de pulmões ou bexiga natatória, cauda heterocerca, intestino com válvula espiral e um clásper pélvico nos machos. A fecundação é interna. Eles podem ser ovíparos ou incubar os jovens internamente, com dependência variável de vitelo ou material nutritivo. Nos tubarões e raias da subclasse Elasmobranchii, cada bolsa branquial abre-se independentemente na superfície corporal. As quimeras da subclasse Holocephali possuem uma dobra opercular que recobre as bolsas branquiais Os tubarões são predadores: as raias são achatadas, vivendo no fundo do mar, onde alimentam-se de moluscos e crustáceos.
A maioria dos peixes atuais é óssea e pertence à classe Osteichthyes. As escamas ósseas continuam na maioria dos casos. O esqueleto interno é, em parte ou quase totalmente, ossificado. Pulmões ou uma bexiga natatória estão presentes. A cauda em geral, é homocerca. A válvula espiral foi perdida na maioria das espécies e cecos pilóricos estão presentes. As brânquias são revestidas por um opérculo. A fecundação é externa e o desenvolvimento é ovíparo na maioria. 0s peixes ósseos ancestrais viviam em água doce sujeitos a estagnação sazonal e seca. 0s pulmões provavelmente evoluíram como um órgão de respiração acessório. 0s peixes pulmonados que permaneceram dulcícolas continuam a ter pulmões. Outros tornaram-se marinhos e os pulmões transformaram-se numa bexiga natatória hidrostática. Muitos desses peixes voltaram à água doce e não perderam a bexiga natatória.
A classe Osteichthyes está dividida em três subclasses. Os Acanthodii, um grupo extinto, apresentava nadadeiras pares com uma base larga, sustentadas por espinhos simples. Os Actinopterygii (percas e espécies assemelhadas) tem nadadeiras pares em forma de abano sustentadas por raios moles. Os Sarcopterygii (peixes pulmonados e crossopterígios) possuem nadadeiras pares lobuladas, sustentadas por um eixo central, carnoso e ósseo. A subclasse Actinopterygii está dividida em três infraclasses: Chondrostei, representada por algumas espécies consideradas relíquias (Polypterus e Acipeonser); Holostei, também representada por algumas espécies relíquias (Lepisosteus e Amia), e Teleostei, que inclui a maioria das espécies atuais.
Durante a evolução dos condóstreos, mais primitivo que os teleósteos, os pulmões transformaram-se numa bexiga natatória, a cauda heterocerca tornou-se homocerca e as escamas ganóides modificaram-se para ciclóides. Os teleósteos, durante o curso de sua evolução, tornaram-se mais hábeis; a nadadeira s original única dividiu-se: as nadadeiras pélvicas deslocaram-se para a frente; espinhos desenvolveram-se na maioria nadadeiras; as escamas mudaram de clclóide para ctenóide e estenderam-se pelo opérculo e cabeça; a bexiga natatória perdeu a conexão com o trato digestivo e a boca tornou-se bastante protrátil. Os teleósteos sofreram uma enorme adversidade adaptativa. Os sarcopterígios são agrupados em duas ordens. Os Dipnoi (peixes pulmonados) possuem o esqueleto ossificado e placas dentárias para esmagar o alimento, constituído de crustáceos e moluscos; três espécies sobrevivem atualmente nas áreas tropicais da América do Sul, África e Austrália. Os crossopterígios têm um esqueleto bastante forte e muitos dentes cônicos. A maioria está extinta, mas o celacanto marinho ainda sobrevive. Os vertebrados terrestres evoluíram a partir dos crossopterígios primitivos dulcícolas.
VERTEBRADOS, PEIXES VERTEBRADOS, PEIXES
Habitats das profundezas
As águas superficiais do oceano são bem iluminadas, bem misturadas, e têm a capacidade de suportar ativamente a fotossíntese de algas. Abaixo desta zona as condições mudam drasticamente. Entre os 200 m e os 1000 m (zona mesopelágica) a luz vai-se gradualmente extinguindo e a temperatura baixa para um termoclima quase permanente entre os 4ºC e 8ºC. O nível de nutrientes, o oxigênio dissolvido e a taxa das correntes também diminui, enquanto a pressão aumenta. Abaixo dos 1000 m (zona batipelágica) as condições são mais uniformes até o fundo ser atingido (zona bentónica profunda), caracterizado pela completa escuridão, temperaturas baixas, poucos nutrientes, nível reduzido de oxigênio dissolvido e elevadas pressões. Este ambiente é o mais extenso habitat aquático na terra. Com a profundidade média dos oceanos de 4000 m, cerca de 98% da sua água é encontrada abaixo dos 100 m e 75% abaixo dos 1000 m.
A vastidão deste ambiente, acoplado com a provável estabilidade ao longo do tempo geológico, levou ao desenvolvimento de uma diversa e quase bizarra ictiofauna, que compreende 11% de todas as espécies de peixes conhecidos. Provavelmente os peixes mais numerosos existentes são as formas pelágicas pequenas (menos de 10 cm), principalmente os “bristlemouths” (Cyclothone). A presença de numerosos e pequenos peixes, dão a impressão de um falso fundo, quando são recebidos os impulsos de um sonar.
Diversidade e distribuição dos peixes de profundidade
O sistema moderno de classificação de seres vivos em geral e de peixes de profundidade em particular data de 1753, quando o sueco Carl Linnaeus, introduziu o seu sistema da nomenclatura binomial.
A descoberta e descrição de novas espécies tem continuado desde então, e de acordo com Cohen (1970), os peixes demersais profundos constituem 6,4% do total de peixes cartilagíneos e ósseos recentemente conhecidos, estando 1280 espécies abaixo dos 200 m.
Estes peixes ocupam áreas muito vastas, mas com condições relativamente estáveis e uniformes, sendo poucos os diferentes nichos disponíveis.
Os peixes oceânicos podem ser divididos em dois grandes grupos, devido à sua ecologia, que é refletida nas suas adaptações morfológicas, afinidades taxonômicas, assim como a sua fauna parasita. As espécies podem ser pelágicas, vivendo em águas abertas, ou demersais, vivendo no fundo (bentônicos), ou imediatamente acima do fundo do oceano (bentopelágicos).
Enquanto que formas epipelágicas são grandes e robustas, como os tubarões ou atuns, os peixes meso ou batipelágicos, são pequenos. A ictiofauna demersal, por seu lado, inclui geralmente formas de maiores dimensões e mais robustas que as pelágicas, como quimeras, tubarões, raias, enguias, tipos de salmão e bacalhau. Em alguns grupos, só algumas espécies ou gêneros ocorrem na profundidade, existindo outros em que, famílias e ordens inteiras, estão restritas a este ambiente.
As observações feitas em peixes demersais não são em grande número, sendo muitos tratamentos e investigações feitos baseados em menos de mil amostras. Muitas espécies foram conhecidas através de pescas de arrasto a grandes profundidades, sendo recentemente utilizados submersíveis para constatar a diversidade e abundância das espécies no seu habitat. Estas técnicas não são muito rigorosas, faltando muitos conhecimentos sobre o seu potencial e seletividade.
Em 1898, Woodward escreveu que as formas vivas ultrapassadas, que não conseguiam competir com as raças vigorosas tiveram que ocupar as águas doces, ou as águas profundas. Esta questão, de onde apareceram as formas mais adaptadas e para onde se retiraram as formas mais antigas, menos adaptadas e mais “fracas”, continua a intrigar os cientistas e a fomentar debates científicos e literatura. Cinqüenta anos mais tarde, Andriyashev (1953), considerou que a adaptação evolutiva dos peixes às profundidades, tinha sido feita por dois grupos que teriam colonizado as profundezas em diferentes alturas.
As formas antigas eram Teleósteos primitivos que teriam habitado todo a variedade de profundidades onde dominaram a fauna demersal, abissal e batipelágica de todo o Oceano. Muitos exibiam adaptações estruturais à vida a grandes profundidades como a proliferação de órgãos luminosos, e modificações nos olhos e na bexiga gasosa.
As outras formas, que ele as chamou de secundárias, seriam derivadas de diversos grupos filogenéticos, e como teriam migrado para as profundezas muito mais tarde que os anteriores, não tinham desenvolvido adaptações externas tão especializadas para aquele ambiente.
Adaptações dos peixes de profundidade
As modificações morfológicas dos peixes das profundezas são o resultado, numa perspectiva evolutiva, das pressões seletivas que este meio foi fornecendo ao longo do tempo. O conjunto de todas essas características, como a coloração, a estrutura mandibular, a musculatura, o posicionamento das barbatanas, e os olhos, fornece-nos, para cada espécie, uma expressão morfológica que define e individualiza os diferentes nichos ecológicos.
O fenômeno de adaptação evolutiva inclui em si outras particularidades que não são tão óbvias, como as diferenças entre as estruturas morfológicas. A adaptação do animal no seu tempo de vida é um desses exemplos, e de modo a se obter uma compreensão generalizada de todo o processo de adaptação é necessário o estudo de vários espécimes em diferentes estágios de vida.
A grande maioria dos peixes que vivem nas profundezas têm um comportamento bentopelágico de modo a minimizar o gasto de energia. Nadam acima do fundo, permanecendo com uma flutuabilidade neutra devido à baixa densidade óssea e à bexiga gasosa. Mesmo espécies que vivem a mais de 5000 m de profundidade, onde a pressão excede as 500 atmosferas, conseguem secretar gases para a bexiga gasosa, através de um sistema de contracorrente que envolve glândulas altamente especializadas.
A sua zona dorsal apresenta uma coloração escura, de modo a se confundirem com o ambiente. Esta coloração tanto pode ser conferida por pigmentação preta, como por pigmentação vermelha, por ser esta uma das primeiras cores do espectro visível a se perder quando luz penetra nas águas oceânicas. Regra geral, os flancos laterais desses peixes são metalizados de modo a refletirem a luz incidente, camuflando-os de todos os ângulos visíveis.
As adaptações mais padronizadas que os peixes, tanto pelágicos como bentônicos, sofreram quando migraram para as profundezas dos oceanos tiveram haver com o gradiente vertical de intensidade de luz.
A rápida diminuição na intensidade de luz com a profundidade resultou num aperfeiçoamento, ou modificação, nos sistemas sensoriais dos peixes bentônicos das profundezas. Surpreendentemente, ao contrário dos peixes pelágicos das profundezas, não ocorreu nenhuma diminuição de complexidade e tamanho dos seus sistemas oculares. Muitos dos teleósteos das profundezas possuem mesmo grandes olhos, com diferentes adaptações, de modo a aumentar a sua sensibilidade e campo visual.
A retenção de luz funcional e substancial, a profundidades abaixo dos 1000 m, por parte dos olhos desses peixes, tem de ser em resposta ao fenômeno da bioluminescência. Muitos dos seres que coabitam nesse ambiente, incluindo invertebrados, possuem estruturas bioquímicas que lhes permitem a emissão de luz. Um dos seus pressupostos será o uso dessas estruturas, associadas aos órgãos de visão, para reconhecimento intra e interespecífico.
Curiosidades:
Entre os cavalos-marinhos, é o macho que dá a luz aos filhotes. A fêmea produz os óvulos e os coloca em um saco de incubação no abdome do macho. Lá, o macho lança os espermatozóides (fecundação externa) e os fertiliza. Nessa bolsa, os embriões se desenvolvem até o vigésimo primeiro dia, quando se dá o "parto".
Podem nascer até 450 ou 600 filhotes de cada vez. Durante o período de gestação, a fêmea visita o macho e os dois realizam rituais de comunicação que os aproximam. Nessa época, eles ficam mais brilhantes e "dançam" ao redor de uma planta marinha, agarrados pelo rabo.
O peixe é riquíssimo em proteínas e fósforo, além de outras substâncias necessárias ao nosso corpo. É um alimento extremamente saudável. Deve ser consumido por todos, desde a infância.
Desde a Pré-História, em qualquer lugar do nosso planeta, o homem vem capturando peixes para se alimentar. A pesca, a venda e a industrialização constituem um ramo muito forte da economia de diversos países como, o Japão, a Noruega, a Rússia, Portugal, peru, etc.
O Brasil possui uma indústria pesqueira em grande desenvolvimento. Mas, como tem um litoral de mais de 9.000 quilômetros, além de muitos rios - maior bacia hidrográfica do mundo -, conta com possibilidades de ser um dos maiores produtores mundiais de pescados e a garantia de alimentação de sua população.
http://www.portalbrasil.net/educacao_seresvivos_vertebrados_peixes.htm
7° série: SISTEMA CIRCULATÓRIO
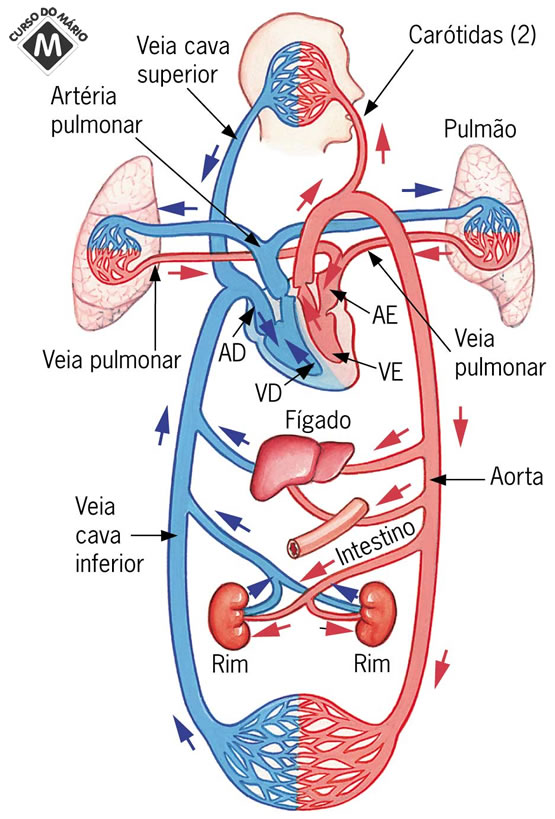


Introdução
O metabolismo requer o constante suprimento de alimento e oxigênio molecular para as células, e o funcionamento das células produz substâncias que devem ser excretadas. A difusão de partículas entre as células não é suficiente para o trânsito das substâncias dentro do organismo. O aparelho circulatório realiza o transporte, entre longas distâncias, de moléculas de um ponto a outro do organismo multicelular.
As partes principais do aparelho circulatório são:
o sangue (composto de um plasma líquido e de células livres), o coração (órgão com paredes musculares que se contraem ritmicamente para impulsionar o sangue através do corpo) e os vasos sangüíneos, onde o sangue circula de modo fechado (o sangue não sai dos vasos sangüíneos). O coração e os vasos sangüíneos são denominados conjuntamente de sistema cardiovascular. Um rápido fluxo de volume de sangue por todas as partes do organismo, através dos vasos sangüíneos, é produzido pelas pressões geradas pela ação bombeadora do coração. A extraordinária arborização dos vasos sangüíneos assegura que todas as células do corpo estejam muito próximas dos menores e mais finos vasos, os capilares.
O sangue e os vasos sangüíneos
A Fisiologia, como ciência experimental, teve início em 1628, quando Willian Harvey demonstrou que o sistema cardiovascular forma um círculo, de maneira que o líquido circulatório é bombeado continuamente desde o coração até um sistema de vasos e retorna ao coração por outro sistema de vasos.
Denominamos veias os vasos que chegam ao coração. As artérias são vasos que saem do coração, dirigindo-se a todas as partes do organismo. É imprópria a denominação "sangue venoso" e "sangue arterial" para nos referirmos aos sangues com alta concentração de dióxido de carbono ou de oxigênio, pois uma veia pode conter sangue com muito ou pouco oxigênio, e o mesmo podemos dizer das artérias, que podem ter sangue com muito ou pouco dióxido de carbono. Portanto, usa-se a denominação sangue carbonado para aquele com alta concentração de gás carbônico, e sangue oxigenado para o sangue que possui expressiva concentração de oxigênio molecular.
O sangue é o líquido circulatório. É composto de um plasma quase incolor onde estão mergulhados elementos celulares: os glóbulos brancos (leucócitos), os glóbulos vermelhos (eritrócitos ou hemácias) e fragmentos celulares (plaquetas ou trombócitos).
O plasma transporta pequenas moléculas alimentícias (aminoácidos, glicose) em solução, metabólitos, secreções internas (hormônios), gases (CO2) e íons. O plasma sangüíneo tem cerca de 92% de água, além de proteínas e cerca de 0,9% de íons inorgânicos (Na+, Cl-). Uma pessoa adulta tem, em média, 5 litros de sangue (cerca de 60% são formados de plasma), portanto, a perda de sangue por hemorragia pode ser fatal.
Elementos celulares do sangue
As hemácias são nucleadas e ovais em todos os vertebrados, com exceção dos mamíferos, nos quais são anucleadas, circulares e bicôncavas.
Os eritrócitos de mamífero são nucleados durante sua formação na medula óssea. O citoplasma da hemácia é preenchido por hemoglobina, um pigmento que tem afinidade com o oxigênio molecular. Como não possui organelas, o metabolismo do eritrócito é limitado; existem as enzimas da glicólise. Em média, há cerca de 4,5 milhões de hemácias por mililitro cúbico de sangue na mulher e 5 milhões no homem. O número total de eritrócitos, num ser humano, é de 30 trilhões. Cada hemácia pode viver 120 dias e fazer 170.000 ciclos de viagem dentro do aparelho circulatório. As hemácias envelhecidas são identificadas pelo seu glicocálix e retiradas de circulação e destruídas pelo baço, de onde grande parte da hemoglobina é passada ao fígado; o pigmento é excretado na bile, e o ferro volta para a medula óssea. Por não terem núcleo, os eritrócitos têm um período de vida limitado.
Os leucócitos têm suas atividades nos vários tecidos do corpo. Os que se encontram no plasma sangüíneo estão, em grande parte, em trânsito de sua fonte (medula óssea, baço, estruturas linfóides) para os tecidos do organismo. Em geral, o período de vida de um leucócito é de 12 a 13 dias.
Os glóbulos brancos podem realizar movimentos amebóides, atravessar o endotélio dos capilares e alcançar os espaços intercelulares dos tecidos. Muitos leucócitos agem como fagócitos, englobando (fagocitando) bactérias que ocorrem em ferimentos; outros produzem anticorpos para defesa imunológica. Nas infecções agudas, como pneumonia, o número de leucócitos sobe de 5.000 a 9.000 (normal) para 20.000 ou 30.000 por mililitro cúbico, com o objetivo de combater a infecção. O pus é a mistura de leucócitos mortos, células dos tecidos e soro sangüíneo.
As plaquetas (ou trombócitos) constituem fundamental elemento do líquido circulatório. São aproximadamente discoidais, anucleadas e muito menores que as hemácias.
No homem, há mais de um trilhão de plaquetas, e cada uma vive de 8 a 10 dias. Quando ocorre uma lesão num vaso sangüíneo, as plaquetas agrupam-se e desintegram-se, liberando a tromboplastina, que inicia o processo de coagulação do sangue.
Funções do sangue
O sangue, nosso líquido circulatório [também existe a linfa], transporta:
a) oxigênio molecular dos pulmões para os tecidos e dióxido de carbono no sentido inverso.
b) água e alimentos obtidos do processo digestivo.
c) alimentos armazenados de um órgão ou tecido para outro, por exemplo, a glicose guardada sob forma de glicogênio.
d) resíduos metabólicos, excesso de água ou íons minerais para os órgãos excretores.
e) hormônios das glândulas onde são produzidos para os tecidos com as células-alvo de sua ação.
f) anticorpos para a defesa do organismo e imunização.
O sangue controla o pH dos tecidos, participando da homeostase, dentro de limites estreitos, por tampões fosfato e bicarbonato. O sangue é ligeiramente alcalino [pH = 7,4].
Capilares e o sistema linfático
Quando o sangue chega no nível dos capilares, realiza a função fundamental do sistema circulatório: a troca de nutrientes e de produtos finais do metabolismo.
Essas trocas ocorrem no líquido intercelular que se localiza entre os capilares e as células. Este líquido é a linfa, um filtrado do plasma, um meio que se origina da filtração de água e solutos através das paredes dos capilares. A saída de plasma acontece na extremidade arterial dos capilares por meio da pressão hidrostática resultante da atividade bombeadora do coração. As proteínas do plasma permanecem nos capilares por causa do seu grande tamanho molecular.
Não existe uma constante perda de plasma do sangue porque a força hidrostática é contrabalançada pela pressão de osmose, que provoca a volta de água aos capilares.
Entre as células e os capilares existem os vasos linfáticos. Esses têm paredes delgadas e com válvulas que impedem o retorno da linfa no seu interior. A maioria dos vasos linfáticos é estrutura tão delicada que não é vista nas preparações anatômicas.
Os vasos linfáticos tornam-se maiores na região do tórax, onde se reúnem para formar o duto torácico, que desemboca no sistema venoso perto do coração. Os vasos linfáticos são a principal via de transporte que os lipídios absorvidos no intestino percorrem para chegar ao sangue. O colesterol dos tecidos alcança o sangue pelos vasos linfáticos. Ao longo do sistema linfático, temos muitos nódulos (gânglios) linfáticos. Neles há a produção de linfócitos. As infecções podem ser acompanhadas de inchação dos nódulos linfáticos.
O coração e o baço
O coração é um órgão muscular, oco, com quatro cavidades (duas aurículas, ou átrios, e dois ventrículos).Esse órgão localiza-se na cavidade torácica e é recoberto por um revestimento fibroso, o pericárdio. As paredes do coração são constituídas principalmente de músculo (o miocárdio).
Coração
( Clique para Ampliar )
O coração recebe sangue das veias e o impulsiona para as artérias. O lado direito impulsiona sangue vindo do corpo em direção aos pulmões. É o que denominamos circuito pulmonar ou pequena circulação. O lado direito bombeia sangue oriundo dos pulmões em direção aos demais órgãos do corpo - é o chamado circuito sistêmico ou grande circulação.
O músculo cardíaco é formado por um tipo especial de fibra muscular estriada. A contração do músculo cardíaco, à semelhança dos outros tipos de células contráteis, é devida à despolarização de sua membrana plasmática. Desde o período embrionário, quando é formado, o coração tem a capacidade de produzir o estímulo para sua própria contração rítmica. Se o coração for retirado e mantido em solução fisiológica, continuará batendo mesmo fora do corpo. A regulação primária da pulsação cardíaca depende de grupos de células cardíacas especializadas. O nódulo sinoatrial (na parte superior da aurícula direita) inicia a contração das aurículas. Logo após, estimula o nódulo atrioventricular (no septo entre as aurículas), que, por sua vez, age sobre um feixe de fibras, o feixe de His (na parede dos ventrículos). Este feixe causa a contração simultânea da musculatura dos ventrículos.
O que ocorreria se todas as diversas fibras musculares cardíacas se contraíssem independentemente, de modo aleatório? Uma conseqüência seria a falta de coordenação no bombeamento do átrios e ventrículos. Com isto, haveria um descontrole do bombeamento de sangue para o organismo. O correto e natural é que as complexas massas musculares que constituem as bombas ventriculares devem se contrair mais ou menos simultaneamente para resultar num eficiente bombeamento. Tal coordenação é resultado por junções celulares que permitem a difusão de um potencial de ação de uma fibra muscular a outra, de modo que a excitação de uma fibra se propaga por todo o coração. Se células cardíacas forem separadas e mantidas em um meio de cultura, a princípio poucas baterão de modo independente. Após alguns dias, as células entram em contato umas com as outras e então todas batem em uníssono.
Há dois tipos de células cardíacas: as que produzem o estímulo para contração e as que, em grande número, formam a massa de tecido que compõe as paredes contráteis do coração. Portanto, algumas células musculares cardíacas são auto-rítmicas, ou seja, são capazes de auto-excitação espontânea. O batimento cardíaco também é controlado pelo sistema nervoso. O nódulo sinoatrial é o marcapasso normal para todo coração. Pode ser estimulado por fibras parassimpáticas (do nervo vago) ou pela presença de acetilcolina. O efeito obtido é a diminuição do ritmo cardíaco. Se a estimulação é realizada por fibras simpáticas ou norepinefrina, o coração tem seu ritmo acelerado.
O trabalho do coração é realizado da seguinte maneira: as aurículas enchem-se de sangue vindo das veias e então se contraem, lançando o sangue para os ventrículos.
Quando estes iniciam sua contração, as válvulas bicúspide (ou mitral) e tricúspide fecham pelo aumento da pressão do sangue e deste modo impedem o retorno do sangue às aurículas. As válvulas semilunares estão ainda fechadas, e a pressão sangüínea nos ventrículos sobe. Quando a pressão nos ventrículos supera a das artérias (os ventrículos se contraem), as válvulas semilunares abrem-se e o sangue é impulsionado para o sistema arterial. Este ciclo se repete durante toda a vida de um indivíduo.
O coração de uma pessoa normal e em repouso bate cerca de 72 vezes por minuto e impele cerca de 60 ml de sangue por ciclo de bombeamento. Durante uma vida de 70 anos, o coração pode contrair-se três bilhões de vezes e impulsionar cerca de duzentos milhões de litros de sangue. Denomina-se débito cardíaco o volume de sangue ejetado por cada ventrículo, por minuto.
A partir do coração, o sangue se movimenta dentro das artérias numa série de "jorros" ou "ondas", percebidas em artérias superficiais (como no pulso ou nas têmporas) como pulsação cardíaca, mais forte quando o coração se contrai (sístole) e mais fraca quando se enche (diástole). O fluxo de retorno de sangue, pelas veias, é praticamente uniforme. As pressões normais para o homem normal, em milímetros de mercúrio, são: artérias, 120/80 (sistólica, diastólica); capilares, 30/10; veias, 10/0. A pressão sangüínea é afetada pelo ritmo dos batimentos cardíacos, constrição dos vasos, entre outros fatores. O médico, auscultando com o estetoscópio, quando ouve os sons de abertura e fechamento das válvulas cardíacas, pode identificar problemas relativos ao aparelho circulatório. O eletrocardiograma é um registro das correntes elétricas produzidas nas fases do trabalho do coração; e também tem a utilidade de auxiliar o diagnóstico pelo médico.
O coração tem seus próprios vasos para supri-lo de oxigênio e nutrientes. Da artéria aorta, partem as artérias coronárias, que percorrem a superfície do órgão e também se irradiam para o interior da massa muscular cardíaca. O sangue volta pelas veias coronárias e entra no seio coronário, que desemboca na aurícula direita. De 7 a 10 por cento do sangue impulsionado para a aorta toma a direção da circulação coronária, e isto evidencia a grande necessidade de nutrientes e oxigênio no metabolismo cardíaco.
Qualquer oclusão dos vasos coronários é sinalizada por dor aguda (angina pectoris) no tórax e braço esquerdo. O entupimento dos vasos coronários por coágulos e placas lipídicas pode ser fatal.
O baço é um órgão que se comunica com os sistemas circulatório e linfático. Age como reservatório, armazenando de um quinto a um terço de células sangüíneas. Produz glóbulos brancos (linfócitos) e destrói hemácias velhas.
Revestimento dos vasos sangüíneos
O coração e todos os vasos são revestidos internamente por um endotélio liso. As paredes da aorta e das artérias de grosso calibre têm camadas espessas de tecido elástico e fibras musculares. As arteríolas têm apenas fibras musculares lisas.
As veias apresentam paredes finas com fibras conjuntivas, mas com pouco tecido muscular.
Ao contrário das artérias, as veias achatam-se quando vazias. As veias têm um grande número de válvulas que auxiliam na manutenção da corrente sangüínea de retorno ao coração (fluxo retrógrado). As válvulas das veias também têm um papel na neutralização dos efeitos da postura ereta do corpo em relação ao fluxo de sangue e à ação da gravidade. No homem, o aparelho circulatório tem papel na regulação da temperatura corporal. Um excesso de calor age sobre um centro nervoso do bulbo, que causa a dilatação dos vasos sangüíneos superficiais da pele, promovendo a dissipação do calor. O resfriamento do ambiente causa a contração desses vasos com o objetivo de evitar a perda de calor.
Percurso da circulação no homem
Se considerarmos uma célula do sangue viajando no nosso corpo, mergulhada no plasma sangüíneo, observamos o seguinte percurso:
O sangue que vem dos diversos órgãos do organismo passa às veias cava inferior e cava superior para adentrar a aurícula direita. Este sangue tem pouco oxigênio e contém dióxido de carbono. Chamamos este sangue de carbonado. Da aurícula direita, passa ao ventrículo direito, atravessando a válvula tricúspide. Como resultado da forte contração do músculo cardíaco (sístole), o sangue atravessa a válvula semilunar e alcança, pelas artérias pulmonares, os pulmões. Dos pulmões, o sangue percorre os numerosos capilares que vascularizam os alvéolos pulmonares. Aí ocorre a troca de dióxido de carbono pelo oxigênio do ar dentro do alvéolo. Essa troca é denominada hematose. Desse modo o sangue é reoxigenado. Agora o sangue segue para vasos maiores e, depois, para as veias pulmonares, que chegam à aurícula esquerda. Pela válvula bicúspide, o sangue chega ao ventrículo esquerdo, onde, por forte contração muscular (sístole), é lançado na artéria aorta, o maior vaso do corpo e de paredes mais resistentes. Por ter que bombear sangue para todos os órgãos, a parede muscular do ventrículo esquerdo é muito espessa e forte. A aorta se divide primeiramente em diversas artérias grandes e de grossas paredes, depois estas se subdividem cada vez mais, de modo a irrigar todas as regiões do nosso organismo. Das artérias, o sangue percorre as artérias microscópicas e os capilares nos tecidos. O sangue volta ao coração pela parte venosa sistêmica: os capilares unem-se para formar vênulas, estas se reúnem em veias, que, ao final, irão encontrar as veias cavas. Em nenhum ponto do coração ou das veias e artérias há mistura de sangue carbonado com sangue oxigenado, a não ser que haja uma comunicação anormal entre as metades funcionais do coração. Também existem outras importantes rotas para o sangue: o sangue arterial, no abdome, entra num sistema de capilares, que irrigam as paredes dos intestinos, onde é absorvido o alimento digerido. O sangue dirige-se, pela veia porta, para o fígado, onde as substâncias nutritivas são armazenadas e metabolizadas. Do fígado, o sangue sai pela veia hepática e chega à veia cava. Outro importante caminho leva o sangue arterial aos rins. Os rins são os principais órgãos que regulam e controlam os níveis de íons e metabólitos (como a uréia). Durante a passagem do sangue pelos rins, o excesso de água e metabólitos (excretas) são eliminados na forma de uma solução aquosa, a urina. Os rins têm um importante papel no controle da homeostase (equilíbrio interno) do nosso corpo.
Patologias do sistema circulatório
A hipertensão (elevada pressão arterial) é definida como uma alta pressão arterial crônica. Esta doença pode ter um resultado final letal, causando insuficiência cardíaca, acidente vascular cerebral (oclusão ou ruptura de um vaso sangüíneo cerebral) ou uma lesão renal.
A aterosclerose é uma enfermidade caracterizada pelo espessamento da parede arterial com células musculares lisas anormais e depósitos de colesterol e outras substâncias. Sabe-se que o fumo, colesterol aumentado, hipertensão, diabete estão associados a essa doença.
Fonte: www.biomania.com.br
7° série: SISTEMA CIRCULATÓRIO
Entrem neste link, site muito legal com animação sobre o sistema circulatório humano.
http://www.aticaeducacional.com.br/htdocs/atividades/sist_circ/
http://www.aticaeducacional.com.br/htdocs/atividades/sist_circ/
8° série - EXERCÍCIOS - Equilibrio das reações químicas
A) C2H6O + O2 --> CO2 + H2O
B) Na2CO3 + HCl --> NaCl + H2O + CO2
C) C6H12O6 --> C2H6O + CO2
D) C4H10 + O2 --> CO2 + H2O
E) FeCl3 + Na2CO3 --> Fe2(CO3)3 + NaCl
F) NH4Cl + Ba(OH)2 --> BaCl2 + NH3 + H2O
G) Ca(OH)2 + H3PO4 --> Ca3(PO4)2 + H2O
H) Fe2(CO3)3 + H2SO4 --> Fe2(SO4)3 + H2O + CO2
I) Na2O + (NH4)2SO4 --> Na2SO4 + H2O + NH3
J) FeS2 + O2 --> Fe2O3 + SO2
K) NH3 + O2 --> NO + H2O
L) KMnO4 + H2SO4--> Mn2O7 + K2SO4 + H2O
M) CS2 + O2 --> CO2 + SO2
N) H3PO4 + CaO --> Ca3(PO4)2 + H2O
O) Na2CO3 + H3PO4 --> Na3PO4 + H2O + CO2
P) KMnO4 --> K2MnO4 + MnO2 + O2
Q) Na + KNO3 --> Na2O + K2O + N2
R) Ni(CO)4 --> Ni + CO
S) CaC2 + H2O --> C2H2 + CaO
B) Na2CO3 + HCl --> NaCl + H2O + CO2
C) C6H12O6 --> C2H6O + CO2
D) C4H10 + O2 --> CO2 + H2O
E) FeCl3 + Na2CO3 --> Fe2(CO3)3 + NaCl
F) NH4Cl + Ba(OH)2 --> BaCl2 + NH3 + H2O
G) Ca(OH)2 + H3PO4 --> Ca3(PO4)2 + H2O
H) Fe2(CO3)3 + H2SO4 --> Fe2(SO4)3 + H2O + CO2
I) Na2O + (NH4)2SO4 --> Na2SO4 + H2O + NH3
J) FeS2 + O2 --> Fe2O3 + SO2
K) NH3 + O2 --> NO + H2O
L) KMnO4 + H2SO4--> Mn2O7 + K2SO4 + H2O
M) CS2 + O2 --> CO2 + SO2
N) H3PO4 + CaO --> Ca3(PO4)2 + H2O
O) Na2CO3 + H3PO4 --> Na3PO4 + H2O + CO2
P) KMnO4 --> K2MnO4 + MnO2 + O2
Q) Na + KNO3 --> Na2O + K2O + N2
R) Ni(CO)4 --> Ni + CO
S) CaC2 + H2O --> C2H2 + CaO
8° série - Equilibrio das reações químicas
Um equilíbrio químico é a situação em que a proporção entre as quantidades de reagentes e produtos em uma reação química se mantém constante ao longo do tempo. Foi estudado pela primeira vez pelo químico francês Claude Louis Berthollet em seu livro Essai de statique chimique de 1803.
Teoricamente, toda a reação química ocorre nos dois sentidos: de reagentes se transformando em produtos e de produtos se transformando de volta em reagentes. Contudo, em certas reações, como a de combustão, virtualmente 100% dos reagentes são convertidos em produtos, e não se observa o contrário ocorrer (ou pelo menos não em escala mensurável); tais reações são chamadas de irreversíveis. Há também uma série de reações nas quais logo que uma certa quantidade de produto(s) é formada, este(s) torna(m) a dar origem ao(s) reagente(s); essas reações possuem o nome de reversíveis. O conceito de equilíbrio químico praticamente restringe-se às reações reversíveis.
Reversibilidade de reações químicas
Um exemplo de reação reversível é a da produção da amônia (NH3), a partir do gás hidrogênio (H2) e do gás nitrogênio (N2) — que faz parte do Processo de Haber:
N2(g) + 3H2(g) --> 2NH3(g)
Nesta reação, quando as moléculas de nitrogênio e as de hidrogênio colidem entre si há uma certa chance da reação entre elas ocorrer, assim como quando moléculas de amônia colidem entre si há uma certa chance de elas se dissociarem e de se reorganizarem em H2 e N2.
No início do processo, quando há apenas uma mistura de hidrogênio e nitrogênio, as chances das moléculas dos reagentes (H2 e N2) colidirem umas com as outras é a máxima de toda a reação, o que fará com que a taxa (ou velocidade) com que a reação ocorre também o seja. Porém à medida com que a reação se processa o número de moléculas de hidrogênio e de nitrogênio diminui, reduzindo dessa forma as chances de elas colidirem entre si e, conseqüêntemente, a velocidade desse sentido da reação. Por outro lado, com o avançar da reação, o número de moléculas de amônia vai aumentando, o que faz com que cresçam as chances de elas colidirem e de se voltar a formar hidrogênio e nitrogênio, elevando assim a velocidade desse sentido da reação. Por fim chegará um momento em que tanto a velocidade de um dos sentidos quanto a do outro serão idênticas, nesse ponto nenhuma das velocidades variará mais (se forem mantidas as condições do sistema onde a reação se processa) e ter-se-á atingido o equilíbrio químico.
Click na imagem para vê-la maior

Teoricamente, toda a reação química ocorre nos dois sentidos: de reagentes se transformando em produtos e de produtos se transformando de volta em reagentes. Contudo, em certas reações, como a de combustão, virtualmente 100% dos reagentes são convertidos em produtos, e não se observa o contrário ocorrer (ou pelo menos não em escala mensurável); tais reações são chamadas de irreversíveis. Há também uma série de reações nas quais logo que uma certa quantidade de produto(s) é formada, este(s) torna(m) a dar origem ao(s) reagente(s); essas reações possuem o nome de reversíveis. O conceito de equilíbrio químico praticamente restringe-se às reações reversíveis.
Reversibilidade de reações químicas
Um exemplo de reação reversível é a da produção da amônia (NH3), a partir do gás hidrogênio (H2) e do gás nitrogênio (N2) — que faz parte do Processo de Haber:
N2(g) + 3H2(g) --> 2NH3(g)
Nesta reação, quando as moléculas de nitrogênio e as de hidrogênio colidem entre si há uma certa chance da reação entre elas ocorrer, assim como quando moléculas de amônia colidem entre si há uma certa chance de elas se dissociarem e de se reorganizarem em H2 e N2.
No início do processo, quando há apenas uma mistura de hidrogênio e nitrogênio, as chances das moléculas dos reagentes (H2 e N2) colidirem umas com as outras é a máxima de toda a reação, o que fará com que a taxa (ou velocidade) com que a reação ocorre também o seja. Porém à medida com que a reação se processa o número de moléculas de hidrogênio e de nitrogênio diminui, reduzindo dessa forma as chances de elas colidirem entre si e, conseqüêntemente, a velocidade desse sentido da reação. Por outro lado, com o avançar da reação, o número de moléculas de amônia vai aumentando, o que faz com que cresçam as chances de elas colidirem e de se voltar a formar hidrogênio e nitrogênio, elevando assim a velocidade desse sentido da reação. Por fim chegará um momento em que tanto a velocidade de um dos sentidos quanto a do outro serão idênticas, nesse ponto nenhuma das velocidades variará mais (se forem mantidas as condições do sistema onde a reação se processa) e ter-se-á atingido o equilíbrio químico.
Click na imagem para vê-la maior

8° série - Reações Químicas
Uma reação química é uma transformação da matéria na qual ocorrem mudanças qualitativas na composição química de uma ou mais substâncias reagentes, resultando em um ou mais produtos. Envolve mudanças relacionadas à mudança nas conectividades entre os átomos ou íons, na geometria das moléculas das espécies reagentes ou ainda na interconversão entre dois tipos de isômeros. Resumidamente, pode-se afirmar que uma reacção química é uma transformação da matéria em que pelo menos uma ligação química é criada ou desfeita.
Um exemplo de uma reação quimica é que (ambos os regentes em solução aquosa):
NaCl + AgNO3 → NaNO3 + AgCl
Nesta reação química, ao passo que o NaNO3 permanece em solução, formou-se uma ligação entre a prata (Ag) e o cloro (Cl) o que resultou em um produto sólido de cloreto de prata (AgCl), pode-se então dizer que houve uma reação química.
Tipos de reações químicas
Tradicionalmente, as reações químicas podem ser classificadas de acordo com o número de reagentes e produtos em cada lado da equação química que representa a reação:
* reações de síntese, composição ou adição;A+b->AB
* reações de análise ou decomposição;AB->A+B
* reações de simples troca ou deslocamento e
* reações de dupla troca
Outra classificação categoriza as reações em dois tipos:
* reações de oxirredução ou reações redox
* as demais reações
Algumas reações de síntese, algumas de análise, todas de simples troca e nenhuma de dupla troca são reações de oxirredução
Um tipo de reação que não encontra paralelo nas classificações acima é a chamada reação de isomerização.
Ainda existem uma série de reações que são estudadas em Química Orgânica, ou seja, sub-classes de reações, tais como : Reações de Halogenação, Reações de Hidrogenação, Reações de Substituição Nucleofílica etc.
Um exemplo de uma reação quimica é que (ambos os regentes em solução aquosa):
NaCl + AgNO3 → NaNO3 + AgCl
Nesta reação química, ao passo que o NaNO3 permanece em solução, formou-se uma ligação entre a prata (Ag) e o cloro (Cl) o que resultou em um produto sólido de cloreto de prata (AgCl), pode-se então dizer que houve uma reação química.
Tipos de reações químicas
Tradicionalmente, as reações químicas podem ser classificadas de acordo com o número de reagentes e produtos em cada lado da equação química que representa a reação:
* reações de síntese, composição ou adição;A+b->AB
* reações de análise ou decomposição;AB->A+B
* reações de simples troca ou deslocamento e
* reações de dupla troca
Outra classificação categoriza as reações em dois tipos:
* reações de oxirredução ou reações redox
* as demais reações
Algumas reações de síntese, algumas de análise, todas de simples troca e nenhuma de dupla troca são reações de oxirredução
Um tipo de reação que não encontra paralelo nas classificações acima é a chamada reação de isomerização.
Ainda existem uma série de reações que são estudadas em Química Orgânica, ou seja, sub-classes de reações, tais como : Reações de Halogenação, Reações de Hidrogenação, Reações de Substituição Nucleofílica etc.
Assinar:
Postagens (Atom)